Xuguang Zhang1,2,
Shouqian Cheng1,2,
Hailong Li1,2,
Xiaopo Zhang2,
Feng Chen1,2,
Youbin Li1,2,
Junqing Zhang1,2 ![]() ,
Yinfeng Tan1,2
,
Yinfeng Tan1,2
For correspondence:- Junqing Zhang Email: jqzhang2011@163.com Tel:+8689866895337
Received: 24 November 2015 Accepted: 14 May 2016 Published: 28 June 2016
Citation: Zhang X, Cheng S, Li H, Zhang X, Chen F, Li Y, et al. Isolation and identification of two galangin metabolites from rat urine and determination of their in vitro hypolipidemic activity. Trop J Pharm Res 2016; 15(6):1235-1241 doi: 10.4314/tjpr.v15i6.16
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the lipid-lowering activity of two metabolites of galangin, namely, galangin-3-O-β-D-glucuronic acid (GG-1) and galangin-7-O-β-D-glucuronic acid (GG-2).
Methods: Female Sprague-Dawley rats were orally administered with galangin. The two metabolites of galangin were isolated from urine sample and purified using Sephadex LH-20 and semi-preparative high performance liquid chromatography (HPLC). The structures of the metabolites were identified by analyzing spectroscopic data. Hypolipidemic activity was evaluated in HepG2 cells. The down- or up-regulation of lipogenic genes was detected using real-time quantitative polymerase chain reaction (qPCR).
Results: Both metabolites of galangin showed hypolipidemic activity. These activities are closely associated with the down-regulation of lipogenic genes such as SREBP-1a, SREBP-1c, and SREBP-2 transcription factors, and the downstream genes such as FAS, ACC, and HMGR were revealed by real-time qPCR data.
Conclusion: The results show that both metabolites possess better lipid-lowering activities than galangin. These hypolipidemic activities are closely associated with inhibiting key genes or proteins that regulated the biosynthesis of both cholesterol and triglycerides.
Introduction
Obesity is becoming a serious threat to human beings [1]. More accumulation of lipids in liver is a key cause for insulin resistance [2]. Therefore, decreasing lipid deposition in liver through inhibition of lipogenesis or stimulation of lipolysis is an effective way for the prevention and treatment of obesity and diabetes [3-4]. Natural products such as cordycepin [5], resveratrol [6], and berberine [7] have showed ability in suppressing lipogenesis and accumulation of fat, and are considered to be the potential candidates in developing new therapeutic agents.
Galangin, a natural flavonoid obtained from Alpinia officinarum, has been found to possess various biological activities [8,9]. Specifically, this compound showed a significant decrease in serum lipids, liver weight, peroxidation of lipids, and accumulation of hepatic triglycerides, which suggested that galangin has the potential for controlling obesity [10]. Interestingly, previous pharmacokinetic studies have found two metabolites of galangin, and are considered as the effective in vivo components [11-12]. However, the limitations in developing new therapeutic agent from galangin are mainly due to its low bioavailability. Therefore, the metabolites obtained from urine samples may be considered as the better candidates. From this approach, two metabolites namely galangin-3-O-β-D-glucuronic acid (GG-1) and galangin-7-O-β-D-glucuronic acid (GG-2) were obtained from the rat urine. In addition, the effects of these two metabolites on lipid accumulation in HepG2 cells and its hypolipidemic mechanisms were investigated to find potential utility in the prevention and treatment of obesity.
Methods
General experimental procedures
HepG2 cells were obtained from the American Type Culture Collection (Manassas, Virginia, USA) and China Union Medical University. 1H, 13C-nuclear magnetic resonance (NMR) experiments were performed on Bruker spectrometer operating at 400 MHz for 1H and 100 MHz for 13C (ietramethylsilane was used as an internal standard). Chemical shifts were expressed in δ ppm referenced to solvent peaks at δH 2.50 and δC 39.6 in dimethyl sulfoxide (DMSO)-d6, and coupling constants were in Hz. Electrospray ionization-mass spectrometry (ESI-MS) was obtained from a Thermo Scientific LTQ-Orbitrap XL instrument (Bremen, Germany). Sephadex LH-20 (GE Health care) was used for column chromatography. High performance liquid chromatography (HPLC) was performed on LC-6AD equipped with an ultra violet (UV) detector of SPD-10A (Shimadzu), and a column of Zorbax SB-phenyl (250 × 9.4mm, 5 µm, Agilent Technologies Co., Ltd). Mixtures of methanol (CH3OH)/water (H2O) were used as an eluent. All solvents used were of analytical grade.
Reagents
Galangin was extracted from Alpinia officinarum in laboratory previously. The chemical structure of galangin was established by analyzing NMR data and compared with literature. The purity of galangin (> 95 %) was determined using HPLC equipped with a UV detector with an Agilent eclipse XDB-C18 column (5µm, 4.6×250 mm). Then, 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) was obtained from Sigma-Aldrich (Saint Louis, Missouri, USA).
Animals
All animal experiments were performed per International Guidelines for Care and Use of Laboratory Animals [20] and approved by the animal ethics committee of Hainan Medical University (reg. no. 201506017/HMU). Female Sprague-Dawley rats (240 to 300 g) were purchased from DongChuang Laboratory Animal Service Department (Changsha, China). Commercial rat chow was available ad libitum except for an overnight fasting period before dosing. All rats were provided with free access to H2O.
Urine collection
Rats were housed separately in rat metabolic cages. Urine samples were collected before and after 0-24 h oral dose of galangin (90 mg/kg). For the oral administration, 30 mg/mL of galangin solution was dissolved in distilled H2O containing 2 % (w/v) tween-80.
Extraction and isolation of two galangin metabolites
The collected urine samples were pooled. Then, 300 mL of pooled urine samples was diluted with 700 mL of distilled H2O to give 1000 mL of solvent. Then, the solvent was partitioned twice with 300 mL of ethyl acetate and n-butanol to give two extracts, respectively. These extracts were concentrated successively under reduced pressure to obtain the ethyl acetate (2.0 g) and n-butanol (5.0 g). The n-butanol extract was separated with Sephadex LH-20 using CH3OH as an eluent to yield six fractions (Fr.1 to Fr.6). Fr.5 (800 mg) was further purified by semi-preparative HPLC with CH3OH:H2O (40:60) as an eluent to give compounds 1 (6.0 mg), and 2 (8.0 mg).
Cell-based lipid accumulation assay
HepG2 cells were maintained in a Dulbecco’s modified eagles medium (DMEM, Gibco, Grand Island, New York, USA) supplemented with 10 % fetal bovine serum (Gibco) and 100 μg/mL of penicillin/streptomycin (Gibco). After reaching 70-80 % confluence, the cells were incubated in DMEM and 100 μM of oleic acid (OA, Sigma-Aldrich, Shanghai, China) for 12 h, then the cells were treated with 10 μM of indicated compounds or marketed antihyperlidemic drug simvastatin (Sigma-Aldrich) in DMEM and 100 μM of OA or with DMEM and 100 μM of OA alone for another 6 h. Subsequently, the cells were subjected to oil-red O staining, total cholesterol and triglycerides were determined as described previously [19]. Each experiment was repeated for three times.
MTT assay
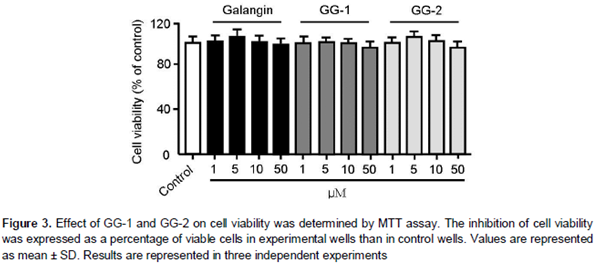
HepG2 cells were cultured in a 24-well plate. After reaching confluence, the cells were incubated for 48 h in presence of GG-1, GG-2, and galangin. Subsequently, the culture medium was removed and replaced with 500 μL of fresh culture medium containing 10 % sterile filtered MTT. After 3 h, the formed insoluble formazan crystals were dissolved in 500 μL of isopropanol per well and absorbance was measured at 570 nm using the 630 nm reading as a reference. The inhibition of growth due to tested compounds was expressed as a percentage of viable cells in experimental wells than in control wells.
Real-time quantitative polymerase chain reaction (qPCR)
The mRNA levels of lipid metabolism-related genes were determined using real-time qPCR. Total RNA extraction, cDNA synthesis, and quantitative PCR assays were all performed as described previously [20]. Samples were cycled 40 times using a fast applied biosystems (ABI)-7500 sequence detector (Foster City, USA). ABI-7500 cycle conditions were as follows: denaturation at 95 °C for 5 min followed by 40 cycles at 95 ºC for 15 s, annealing at 60 ºC for 30 s, and holding at 72 ºC for 30 s. Cycle threshold was calculated under default settings of real-time sequence detection software (Applied Biosystems). Three independent biological replicates were analyzed to check the reproducibility of the data.
Statistical analysis
Data are presented as mean ± standard deviation. One-way analysis of variance was used to determine the significant difference between both groups. Modified Student’s t-test with the Bonferroni correction was used to compare the difference between individual groups. P < 0.05 was considered as statistically significant.
Results
Spectral data for GG-1, GG-2, and galangin
GG-1: ESI-MS m/z 444.9 [M-H]-; 1H-NMR (400 MHz, DMSO-d6) δ: 12.38 (1H, br.s, 5-OH), 8.13 (2H, d, J=7.5 Hz, H-2', 6'), 7.52 (3H, m, H-3', 4', 5'), 6.26 (1H, br.s, H-8), 6.08 (1H, br.s, H-6), and 5.54 (1H, d, J=7.2 Hz, H-1"); and 13C-NMR (100 MHz, DMSO-d6) δ: 178.1 (C-4), 172.2 (C-6"), 163.0 (C-7), 162.6 (C-5), 157.0 (C-9), 156.3 (C-2), 134.5 (C-3), 131.1 (C-1'), 130.9 (C-4'), 129.6 (C-2', 6'), 128.6 (C-3', 5'), 104.3 (C-10), 99.2 (C-6), 94.3 (C-8), 76.7 (C-3"), 74.8 (C-5"), 74.3 (C-2"), and 72.5 (C-4").
GG-2: ESI-MS m/z 444.9 [M-H]-; 1H-NMR (400 MHz, DMSO-d6) δ: 12.38 (1H, br.s, 5-OH), 8.10 (2H, d, J=7.5 Hz, H-2', 6'), 7.52 (3H, m, H-3', 4', 5'), 6.85 (1H, br.s, H-8'), 6.44 (1H, br.s, H-6'), and 5.13 (1H, d, J=7.2 Hz, H-1"); and 13C-NMR (100 MHz, DMSO-d6) δ: 178.1 (C-4), 172.1 (C-6"), 162.6 (C-5), 160.8 (C-7), 156.5 (C-9), 146.8 (C-2), 132.1 (C-3), 131.4 (C-1'), 130.6 (C-4'), 129.0 (C-2', 6'), 128.1 (C-3', 5'), 105.3 (C-10), 99.4 (C-6), 94.8 (C-8), 76.7 (C-3"), 74.7 (C-5"), 73.3 (C-2"), and 72.3 (C-4").
Galangin: ESI-MS m/z 268.9 [M-H]-; 1H-NMR (400 MHz, DMSO-d6) δ: 12.36 (1H, br.s, 5-OH), 8.14 (2H, m, J=7.5 Hz, H-4'), 7.50-7.55 (3H, m, H-3', 4', 5'), 6.45 (1H, br.s, H-8'), and 6.20 (1H, br.s, H-6'); and 13C-NMR (100 MHz, DMSO-d6) δ: 176.3 (C-4), 164.3 (C-7), 160.8 (C-5), 156.5 (C-9), 145.8 (C-2), 137.1 (C-3), 131.4 (C-1'), 130.0 (C-4'), 128.6 (C-2', 6'), 127.6 (C-3', 5'), 103.3 (C-10), 98.4 (C-6), and 93.6 (C-8).
Characteristic features of the two galangin metabolites
Compound 1 was obtained as a brown powder. The spectral data of 1H-NMR were resonated at δ: 12.38 (1H, br.s, 5-OH), 8.13 (2H, d, J=7.5 Hz, H-2', 6'), 7.52 (3H, m, H-3', 4', 5'), 6.26 (1H, br.s, H-8), and 6.08 (1H, br.s, H-6) indicated compound 1 as a flavonoid. The NMR spectral data of compound 1 were similar to that of galangin. Further analysis of NMR data showed that a glucuronic acid group existed in compound 1. Comparing the 13C-NMR data of compound 1 with galangin revealed the downfield shifts of C-2 and C-4 were +10.6 and 1.8, respectively, whereas the upfield shift of C-3 (-1.6 ppm) indicating the glucuronic acid group was located at C-3. Therefore, compound 1 was identified as GG-1.
Compound 2 was also obtained as a brown powder. The spectral data of NMR and MS were identical to compound 1. The connection of glucuronic acid group to the agylcone was also determined in a same manner. Comparing the 13C-NMR data of compound 2 with galangin revealed the downfield shifts of C-6 and C-8 were +1.2 and 1.1, respectively, whereas the upfield shift of C-7 (-3.4 ppm) indicating the glucuronic acid group was located at C-7. Therefore, compound 2 was identified as GG-2.
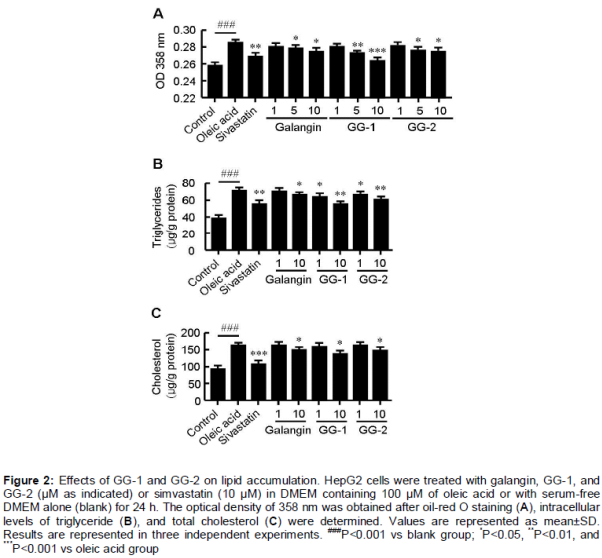
GG-1 and GG-2 inhibit lipid accumulation in HepG2 cells
To evaluate the effect of GG-1 and GG-2 on lipid metabolism, OA-elicited neutral lipid accumulation in HepG2 cells was used. The intracellular lipid content was determined by oil-red O staining and specific kits for accumulation of lipid, total cholesterol, and triglycerides. Supplementation with OA significantly increased accumulation of lipid in HepG2 cells (). Treatment with GG-1 and GG-2 decreased OA-elicited neutral lipid accumulation (A) as well as intracellular contents of triglyceride (B) and total cholesterol (C) in a dose-dependent manner. The inhibitory efficiency of both compounds was higher than that of galangin. The MTT assay showed that the inhibitory effect of GG-1 and GG-2 on lipid metabolism was independent of its cytotoxic effects on HepG2 cells ().
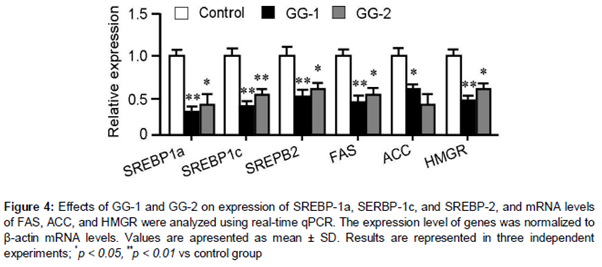
GG-1 and GG-2 decreased transcription of lipogenesis-related transcription factors and its targeted genes
Real-time qPCR showed that treatment with 10 μM of GG-1 and GG-2 significantly decreased the expression of lipogenic genes such as SREBP-1a, SREBP-1c, and SREBP-2 transcription factors and its downstream genes such as FAS, ACC, and HMGR. GG-1 is more potent in regulation of these lipogenic genes than GG-2 ().
Discussion
Herbal medicines have been used in the treatment of obesity for past 100 decades [21,22]. Natural compounds with interesting structures and lipid regulating activities have attracted numerous attentions from chemists and pharmacologists [23]. Statins are the representative compound originated from natural sources with antihyperlipidemic activity [24]. Other compounds such as cordycepin, resveratrol, and berberine are deemed as the effective regulators of lipid metabolism. Galangin has also been proved to have the ability in lowering the lipid levels in liver and blood [10].
However, most natural products are not used as a drug when isolated from natural sources [25]. There are many factors influencing the drug ability. Among all, low biological availability is considered to be the most important factor [26]. Many methods are used to resolve this problem. Finding metabolites from natural products of biological samples have proved to be an effective process in drug discovery [27,28].
Previous studies have found that GG-1 and GG-2 are the main metabolites of blood and urine samples when rats were orally administered with galangin. Therefore, GG-1 and GG-2 are considered to be the active in vivo constituents. To verify this hypothesis, the lipid lowering activity of these metabolites has been investigated. The results have demonstrated that GG-1 and GG-2 possess lipid lowering activity. Significantly, these two metabolites showed better hypolipidemic activities than galangin. This may be attributed to the presence of glucuronic acid group in the carbon skeleton. The potential mechanisms have also been investigated and are found to be closely associated with inhibiting key genes or proteins that regulated the biosynthesis of both cholesterol and triglycerides.
Conclusion
The findings of this study indicate that both metabolites possess better lipid-lowering activities than galangin. These hypolipidemic activities are closely associated with inhibiting key genes or proteins that regulate the biosynthesis of both cholesterol and triglycerides.
Declarations
Acknowledgement
References
Archives


News Updates